苯丙氨酸作为蛋白质的基本构架,是所有生命的基本单元,更是重要神经递质合成的前体物质。迄今为止,人们发现植物中苯丙氨酸的合成途径仅有两条。一是植物中广泛存在的主要合成途径;二是仅在少数植物如松树(Pinus pinaster)、水稻(Oryza sativa)、拟南芥(Arabidopsis thaliana)、矮牵牛(Petunia hybrid)中发现的补偿途径。其中,阿罗酸和苯丙酮酸分别是植物苯丙氨酸主要合成途径和补偿合成途径的重要中间物质。此前,我们未见有关水果类植物存在该补偿合成途径的报道。
三叶木通作为第一个基于游离氨基酸含量而开发的食疗保健水果,富含游离氨基酸(尤其是芳香氨基酸)和各种次生代谢物(如黄酮类化合物)等与苯丙氨酸生物合成途径密切相关的营养成分,以其独特的营养价值在市场上备受喜爱。然而,植物苯丙氨酸合成途径牵涉调控网络复杂,且其在植物中的分布范围仍不清楚。近日,3200威尼斯vip罗培高教授团队在期刊International Journal of Biological Macromolecules发表研究论文《Arogenate dehydratase isoforms strategically deregulate phenylalanine biosynthesis inAkebia trifoliata》,研究在三叶木通苯丙氨酸合成调控上取得突破性进展。

该研究从三叶木通基因组中鉴定出8个ADT家族成员(AktADT1-8),进一步利用生物化学、遗传学等方法分析了阿罗酸脱水酶(ArogenateDehydratase, ADT)家族的结构特征、进化经历、理化特性,并系统的研究了其酶学特性及其在苯丙氨酸合成中的管制作用。
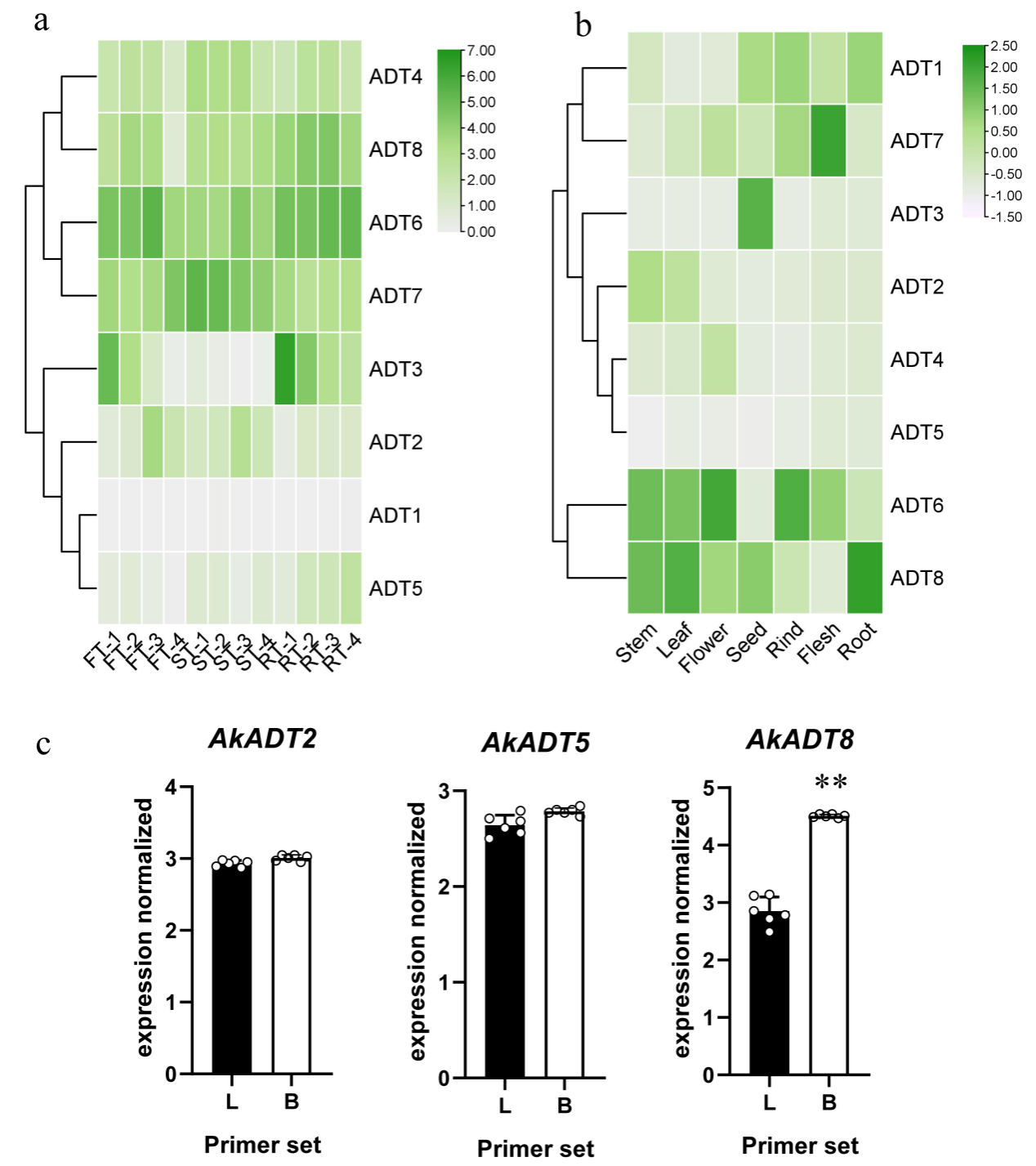
转绿组数据和qRT-PCR结果均显示,AktADTs基因呈现出明显的组织与时间特异性表达,这可能与其功能密切相关。序列比对发现AktADT2、AktADT5和AktADT8的开放阅读框内存在第二个起始密码子,分别位于第一个起始密码子下游的第133、54和177个氨基酸;利用植物启动子数据库对转录起始位点进行预测,结果表明仅有AktADT8存在2个转录起始点,随后利用同一基因不同区域的特异引物qRT-PCR分析进一步证实了这一事实。
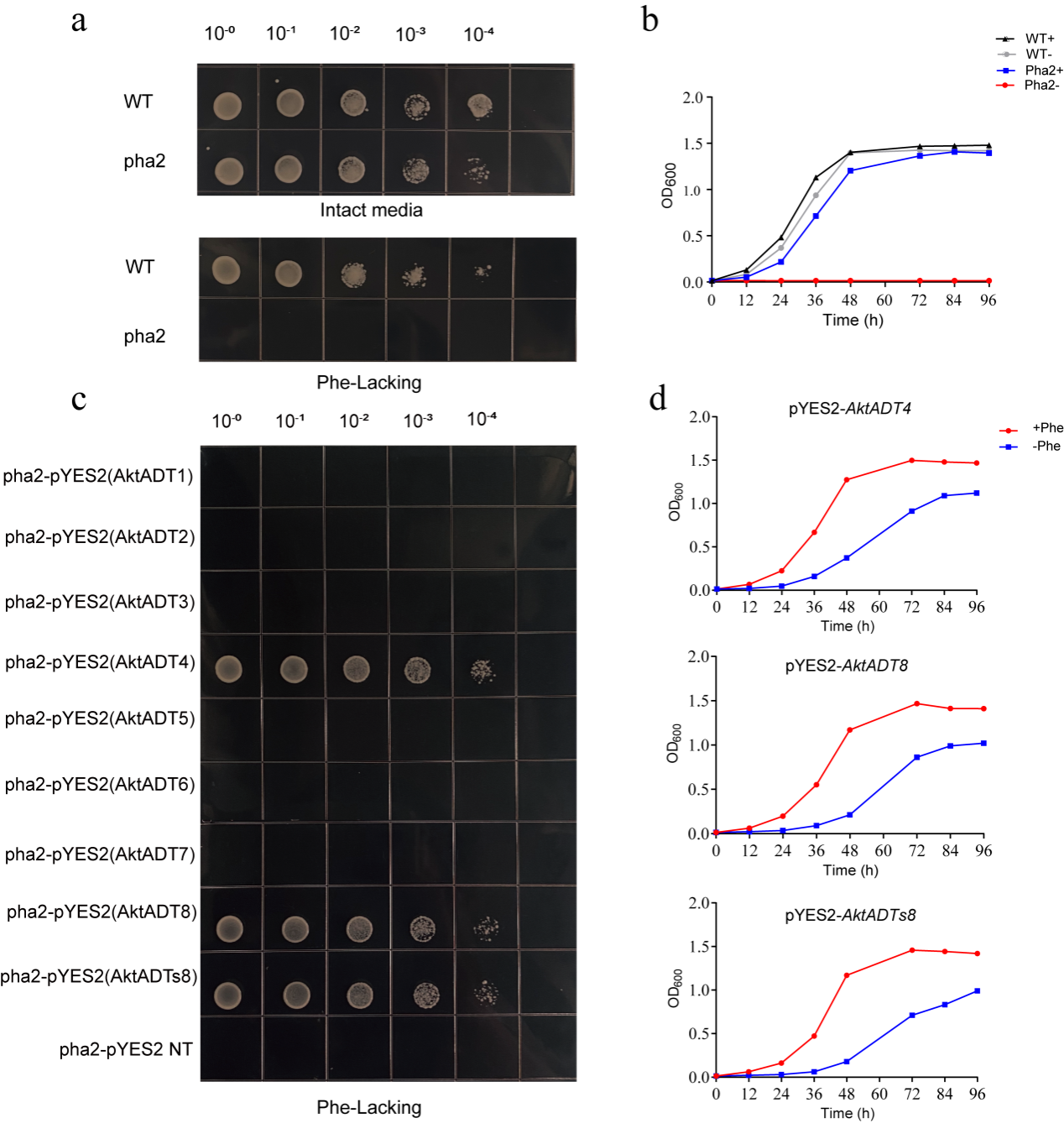
研究证实了AktADT1、4、7、8及s8具有底物混杂性可利预苯酸作为底物行使PDT功能,进一步在酵母pha2敲除株中转化诱导表达,AktADT4、AktADT8和AktADTs8能在酵母体内行使PDT功能从而恢复酵母敲除株pha2的生长表型。且系统发育分析表明,PAC结构域的两个基本残基组合(Val314/Ala317或Ala314/Val317)对ADT的兼性PDT活性至关重要。
研究表明AktADTs具有不同程度的苯丙氨酸反馈抑制敏感性(IC50),AktADT5对苯丙氨酸的反馈抑制最不敏感,其次是AktADT8,而AktADT7反馈抑制最为敏感。在烟草细胞中瞬时表达AktADTs-GFP融合蛋白,对AktADTs及截短转录本AktADTs8进行了亚细胞定位,结果表明AktADTs均定位于叶绿体,而AktADTs8定位于细胞质中。进一步观察发现AktADT4和8定位于叶绿体基质微管结构,其余AktADTs定位于叶绿体基质。论文证实了三叶木通具有完整的胞质定位苯丙酮酸途径催化链,为苯丙氨酸合成及下游次生代谢调控提供了新视角。研究揭示了AktADTs调控苯丙氨酸积累的分子机制,为未来利用基因和代谢工程,提高三叶木通苯丙氨酸含量提供了理论基础。
